This post is about what indirect genetic effects are, and where they came from. It’s about an interesting evolutionary process, and a bit of sleuthing into the depths of a library to find a dusty old book.
A direct genetic effect is what you think of when the classic “nature vs nurture” debate comes up. It is the genes in an individual that affect some aspect of its phenotype (how it looks, how it acts, what its physiology is like etc). For any aspect of a population’s phenotypes, e.g. peoples’ heights, given the right information we can work out what % of the variation is due to direct genetic effects (i.e. nature), and the remaining portion is assumed to be caused by the environment (as well as “non-additive” genetic effects like dominance, but we won’t worry too much about those effects here). With me so far? Excellent!
Now, an indirect genetic effect occurs when two (or more) individuals interact with each other, such as when two stags fight each other, or a mother cow feeds its offspring milk. In this case, some aspect of an individual’s phenotype, such as how heavy the calf is or how successful the stag is, depends on the actions of the individual it is interacting with, in the case of the calf this is the mother, while in the case of the stag it is the rival stag. As the actions of the mother (and the rival stag) are in part determined by its genes, then some of the effect of the mother (and the rival stag) on the calf (and our focal stag) stem from the genes of the interacting partner. So the genes of the mother cow for milk production influence the weight of the calf, while the genes for fighting in the rival stag influence how successful our focal stag.
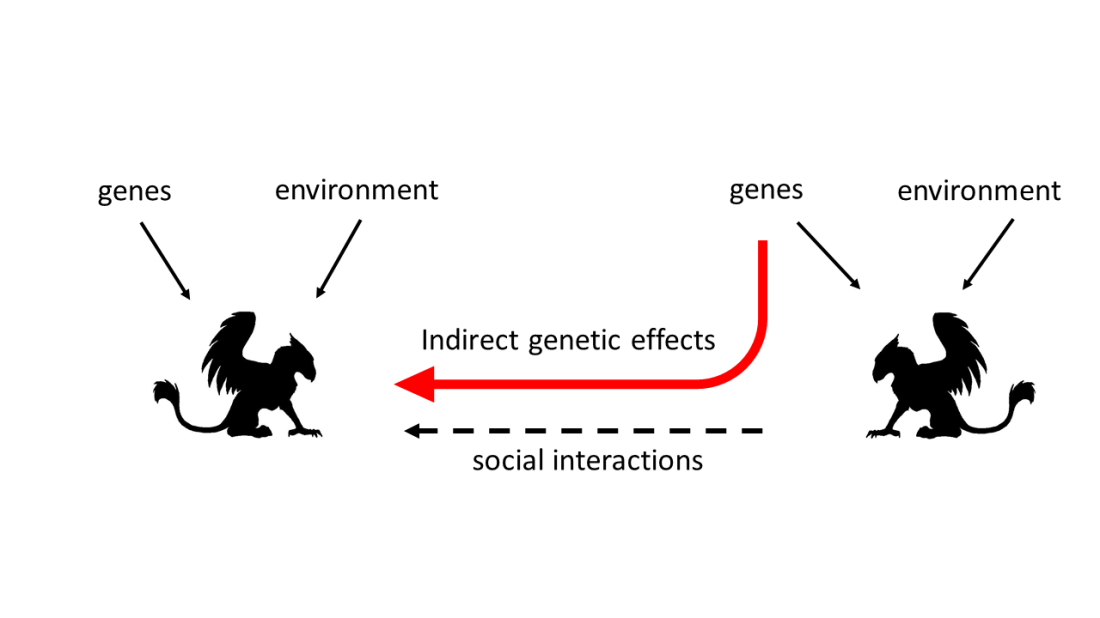
So why are these interesting? Aside from allowing us to say things like “The environment is made of genes” (!!!!) ? Well these lil’ indirect genetic effects (IGEs to the cool kids) can profoundly change how populations evolve. The key thing is how IGEs covary (or are correlated) with direct genetic effects (DGEs hereafter, as we too wish to be cool).
If IGEs and DGEs are positively correlated, then individuals that have genes to (for example) grow fast might also have genes to produce lots of milk, and so cause their offspring to grow fast as well (above and beyond the fact that the offspring have inherited the genes for fast growth directly). This means that if fast growing individuals do better and have more offspring, the next generation will be full of individuals that both grow fast and cause their offspring to grow fast (thanks to their boosted milk production). Therefore, the average growth rate gets a double whammy, and increases especially quickly. So a positive DGE-IGE correlation speeds up evolution. But a negative correlation? What if fast growing mothers are in fact the worst milk producers (and this is all due, at least in part, to genes)? Well if fast growing still leads to more offspring, then the next generation will have more fast growing, but less-milk producing individuals. Therefore, the average growth rate will in fact not increase much at all, as each individual gets less milk during development. In fact, if these fast growing cows are really bad at producing milk, the average growth rate in the population might even decrease, despite fast growth still being associated with increased fitness. So a negative DGE-IGE correlation can reduce, remove, or even reverse any evolutionary change!!

I will add here the only empirical evidence of a reversed response to selection in the presence of IGEs is (probably) that of Wade (1976), who found that populations of flour beetles under selection for increased reproduction actually decreased their reproduction over generations (although Wade did not discuss their results in terms of IGEs). Sometimes when people say a reversed response to selection is possible they point to Goodnight (1985) or a Muir paper (typically Muir 1996 or 2005), but as far as I can tell these studies observed responses to individual selection that were not different from zero. But anyway, still way cool, as they had no response to selection despite strong selection on a heritable trait. So IGEs can fundamentally change evolution.
So, given they are so interesting and potentially important, who first discovered them? Who gets the credit/glory?*
Well, the origin of the name is easy. Moore et al (1997) coined the phrase “indirect genetic effects”, and it has bubbled along from there. But the actual idea (specifically that an individual’s phenotype is dependent on the genes of others, and this can influence evolutionary change)? Well that too seems fairly unequivocal. Most point to Griffing (1967) as the first to realise that the response to selection depends on how animals’ genes influence each other, and that incorporating what he called “associative effects” into the predicted response to selection was critical for animal breeders. So, well done B. Griffing. It may have taken 30 years for the ball to really get rolling, but you gave it the first push.

I’ll note quickly here that R A Fisher (1930) may have stumbled across, or at least wandered past, this idea when he talked about “environmental deterioration” and the evolution of more competitive environments. But we aren’t quite sure what he meant with his fundamental theorem still, so I don’t think he can be given the credit for planting the seed of IGEs.
All well and good and case closed? Well, not quite. While perusing the literature, I stumbled across a reference to “Social genetics”, a review article by J P Scott in 1977. In there, he states “If the behavior of only one individual is measured, it should reflect not only the effects of his own genes but also those of the other individual involved in the relationship (Scott and Fuller, 1965).” my emphasis. Two things to note there:
First, Scott is definitely discussing the idea of IGEs, and appreciates their substantial importance for understanding social behaviour.
Secondly, and hopefully what you’re wondering about, DID SCOTT AND FULLER (1965) ACTUALLY GET THERE FIRST!?!? Did Scott and Fuller (1965) beat Griffing (1967) to the punch, and, even if they didn’t influence the field widely, first appreciate that the environment contains genes? To find out, I had to find this publication.
Turns out, its a book: “Genetics and the Social Behavior of the Dog”. Luckily, my university (the University of Guelph) has a long history in the study of domesticated animals, and a copy of this tome was kept in the dusty reaches of the library. Pretty soon, a creaking hardbound book was in my hands.

Turns out its a book on a huge (13 year) study on dog behaviour, how it depends on genes, experiences and the environment, and how dogs socially interact with each other, and so on. It contains some nice gems, such as:

But enough pearly insights into misguided selection of puppies by humans, do they expose IGEs to the world? Well, I think they did. It took a bit of searching, but eventually I found the following section:
Part IV General Implications
“The behaviour of an individual in a social relationship depends not only upon his own heredity and past environment (experiences) but also on those of the other individual”
Accompanying that statement is this figure:
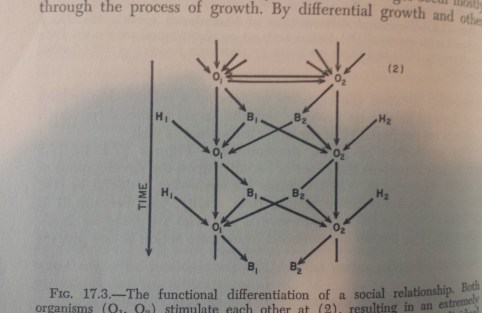
Which illustrates their model for the development of an organism (“O”, with different individuals indicated by numbers as subscripts) over time. O is influenced by its own genes (“H”, for heredity) which, combined with environmental effects which are not depicted in this figure, give its behaviours (“B”). These behaviours feed back onto the organism’s development (B1 leads to O1), but crucially also influence the development of another organism (B1 leads to O2).
So here Scott and Fuller clearly consider the role of the genes of individuals in shaping the phenotypes of others. This is exactly what IGEs are, so I do think they got there first.
However, while they do go on to discuss whether social interactions will speed up or slow down evolution, they do not discuss how DGEs and IGEs might be correlated, and couch none of their discussion in terms of quantitative genetics and the response to selection. This means they do not make the same kind of conclusions about the evolutionary importance of IGEs that Griffing and Moore et al. did later on. Finally, whoever read this book did not pick up on these particular conclusions, as far as I am aware the work of Scott and Fuller does not feature at all in most discussions of the quantitative genetics of social behaviour (although Moore et al. do refer to a Fuller and Hahn (1976) paper which discusses methodological issues surrounding the study of the genetics of social behaviour). So maybe they were a bit ahead of their time, or just didn’t get their work read by the people who would have best made use of it. Ultimately that fell to Moore et al., some 32 years later.
I want to make it clear I do not think Moore et al, as well as Griffing, should not get the credit for IGEs, nor do I think they got the idea from Scott and Fuller and failed to credit them. I simply think that the idea seems to have arisen in more than one research group, and only one of these instances lead to a more general change in our understanding of how evolution works.
What can we take away from this diversion to the far corners of a library?
- Multiple people can have a similar idea, but only one of them may be responsible for its spread
- It takes time for an idea to be fully appreciated
- Old books contain some forgotten gems
* how interesting you find this question probably comes down to personal taste. Maybe you think it doesn’t matter who came up with the idea , just that the idea exists at all. Fair enough, I’m not going to argue with that viewpoint. But I personally find it interesting tracing these threads back through papers to individuals, so that is what I have done here.
References
Fisher, R. A. 1930. The Genetical Theory of Natural Selection. Oxford University Press.
Goodnight, C. J. 1985. The influence of environmental variation on group and individual selection in a cress. Evolution. 39:545–558.
Griffing, B. 1967. Selection in reference to biological groups. I. Individual and group selection applied to populations of unordered groups. Aust. J. Biol. Sci. 20:127–39.
Moore, A. J., E. D. I. Brodie, and J. B. Wolf. 1997. Interacting phenotypes and the evolutionary process: I. Direct and indirect genetic effects of social interactions. Evolution. 51:1352–1362.
Muir, W. M. 1996. Group selection for adaptation to multiple-hen cages: selection program and direct responses. Poult. Sci. 75:447–58.
Muir, W. M. 2005. Incorporation of competitive effects in forest tree or animal breeding programs. Genetics 170:1247–1259.
Scott, J. P. 1977. Social genetics. Behav. Genet. 7:327–346.
Scott, J. P., and J. L. Fuller. 1965. Genetics and the Social Behavior of the Dog. 1st ed. University of Chicago Press, Chicago.
Wade, M. J. 1976. Group selection among laboratory populations of Tribolium. Proc. Natl. Acad. Sci. 73:4604–4607.
